
Структура нуклеиновых кислот нуклеотиды их разновидности. Нуклеиновые кислоты. Спирализация молекулы. Явление комплементарности
Более ста лет назад (в 1869 году) Фридрих Мишер, исследуя клетки гноя, выделил из ядер этих клеток новый тип химических соединений, которые он в совокупности назвал "нуклеином". Эти вещества, позднее получившие название нуклеиновых кислот, обладали кислотными свойствами, были необычайно богаты фосфором и содержали также углерод, кислород, водород и азот. Последующее изучение их показало, что существует два типа нуклеиновых кислот: дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК), которые являются составной частью сложных белков - нуклеопротеинов, содержащихся во всех клетках животных, бактерий, вирусов, растений.
Нуклеопротеины [соответственно, дезоксирибонуклеопротеины (ДНП) и рибонуклеопротеины (РНП)] отличаются друг от друга по составу, размерам и физико-химическим свойствам. Названия нуклеопротеинов отражают только природу углеводного компонента (пентозы), входящего в состав нуклеиновых кислот. У РНП углевод представлен рибозой, у ДНП - дезоксирибозой. Название "нуклеопротеины" связано с названием ядра клетки, где они впервые и были обнаружены. Однако в настоящее время установлено, что ДНП и РНП содержатся и в других субклеточных структурах. При этом ДНП преимущественно локализованы в ядре, а РНП - в цитоплазме. В то же время ДНП открыты в митохондриях, а в ядрах и ядрышках обнаружены также высокомолекулярные РНП.
| Отличия между ДНК и РНК | ||
| Показатели | ДНК | РНК |
| Местоположение | ядро клетки, в составе хроматина, немного в митохондриях (0,2% от всей ДНК) | во всех частях |
| Сахар (пентоза) | Дезоксирибоза | Рибоза |
| Азотистые основания | Аденин, Гуанин, Цитозин, Тимин |
Аденин, Гуанин, Цитозин, Урацил |
| Количество цепей в молекуле | 99,99% - двойная спираль, 0,01% одноцепочечная | 99,99% - одноцепочечная, 0,01% двухцепочечная |
| Форма молекулы | Все одноцепочечные - кольцевые. Большинство двухцепочечных - линейные, часть - кольцевые. |
Линейные молекулы |
Химический состав нуклеиновых кислот
Выделение нуклеиновых кислот из комплекса их с белками и последующий их полный гидролиз позволил определить химический состав нуклеиновых кислот. Так, при полном гидролизе в гидролизате были обнаружены пуриновые и пиримидиновые основания, углеводы (рибоза и дезоксирибоза) и фосфорная кислота.
Азотистые основания (N-основания)
В основе структуры пуриновых и пиримидиновых оснований лежат два ароматических гетероциклических соединения - пурин и пиримидин. Молекула перимидина содержит один гетероцикл. Молекула пурина состоит из двух конденсированных колец: пиримидина и имидазола.
Обрати внимание! Нумерация атомов в ароматическом кольце азотистых оснований осуществляется арабскими цифрами без штриха [ " ]. Символ [ " ] (произносится как "штрих" или "прим") показывает, что соответствующий номер нумерует атомы пентозного кольца, например 1" (см ниже).
В составе нуклеиновых кислот встречаются три главных пиримидиновых основания: цитозин (Ц), урацил (У) и тимин (Т):

и два пуриновых - аденин (А) и гуанин (Г)

Одним из важных свойств азотистых оснований (содержащих оксигруппы) является возможность их существования в двух таутомерных формах, в частности лактим- и лактамной формах, в зависимости от значения pH среды. Таутомерные превращения можно представить на примере урацила.

Оказалось, что в составе нуклеиновых кислот все оксипроизводные пуринов и пиримидинов находятся в лактамной форме.
Помимо главных оснований, в составе нуклеиновых кислот открыты редкие (минорные) азотистые основания. Минорные основания встречаются преимущественно в транспортных РНК, где их список приближается к 50, в следовых количествах в рибосомальных РНК и в составе ДНК. В транспортных РНК на долю минорных оснований приходится до 10% всех нуклеотидов, что имеет, очевидно, важный физиологический смысл (защита молекулы РНК от действия гидролитических ферментов). К минорным основаниям относятся дополнительно метилированные пуриновые и пиримидиновые основания, например, 2-метиладенин, 1-метилгуанин, 5-метилцитозин, 5-оксиметилцитозин и др.


Углеводы
Углеводы (пентозы) в нуклеиновых кислотах представлены рибозой и 2-дезоксирибозой, которые находятся в β-D-рибофуранозной форме (формулы слева).
В составе некоторых фаговых ДНК обнаружена молекула глюкозы, которая соединяется гликозидной связью с 5-оксиметилцитозином.
Конформация углеводного цикла (пентозы)
Для углеводного цикла (пентозы) нуклеиновых кислот плоская конформация, когда атомы углерода С1", С2", С3", С4" и гетероатом кислорода находятся в одной плоскости, - энергетически невыгодна. Среди многочисленных теоретически возможных конформаций этих остатков в полинуклеотидах реализуются только две: либо С2"-эндоконформации, либо С3"-эндоконформации. Эти конформации возникают при вращении вокруг связи С4", которое приводит к такому искажению кольца, при котором один из атомов пентозы (пятичленного фуранозного кольца) оказывается вне плоскости создаваемой четырьмя другими атомами. Такая конформация представляет собой эндо- или экзо- структуру, в зависимости от того располагается ли данный атом на той же стороне плоскости, что и С5" или на противоположной стороне.
Вещества, в которых азотистые основания соединены с пентозой, называются нуклеозидами (рис.2).

Нуклеозиды относятся к N-гликозидам. У них пиримидиновые азотистые основания (один гетероцикл) соединяются с пентозой гликозидной связью через N-1, пуриновые через N-9. В зависимости от типа пентозы различают два вида нуклеозидов - дезоксирибонуклеозиды, содержащие 2-дезоксирибозу, и рибонуклеозиды, содержащие рибозу.
Дезоксирибонуклеозиды входят только в ДНК, а рибонуклеозиды - только в РНК. Пиримидиновые и пуриновые нуклеозиды содержат соответствующие азотистые основания:

Кроме главных встречаются минорные нуклеозиды, в которые входят минорные азотистые основания. Больше всего минорных нуклеозидов содержится в тРНК. Наиболее распространенными минорными нуклеозидами, входящими во все тРНК, являются дигидроуридин, псевдоуридин (обозначаемый сокращенно значком Ψ) и риботимидин. В псевдоуридине отсутствует обычная N-гликозидная связь. В нем атом С-1 рибозы соединен с атомом С-5 урацила.
Вследствие стерических причин пуриновые основания в составе пуриновых нуклеотидов в ДНК могут принимать только две стерически доступные конформации относительно остатка дезоксирибозы, обозначаемые как син-конформации и анти-конформации.
В то же время пиримидиновые основания пиримидиновых нуклеотидов присутствуют в ДНК в виде анти-конформеров, что связано со стерическими несоответствиями, возникающими между углеводной частью нуклеотида и карбонильным кислородом в С-2 положении пиримидина. В силу этого пиримидиновые основания приобретают, главным образом, анти-конформацию (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004).
Нуклеотиды представляют собой соединения соответствующего типа нуклеозида с фосфорной кислотой. Они также делятся на рибонуклеотиды, содержащие рибозу, и дезоксирибонуклеотиды, содержащие 2-дезоксирибозу. Название нуклеотида происходит от вида азотистого основания и количества остатков фосфорной кислоты. Если содержится один остаток фосфорной кислоты - нуклеозид монофосфат (к примеру дAMФ - дезоксиаденозин монофосфат), два остатка – нуклеозид дифосфат (к примеру дAДФ - дезоксиаденозин дифосфат), три остатка – нуклеозид трифосфат (к примеру дAТФ - дезоксиаденозин трифосфат). Остатки фосфорной кислоты присоединяются к 5"-углероду дезоксирибозы и обозначены α, β, γ.
Ниже приведено строение адениловых нуклеотидов.
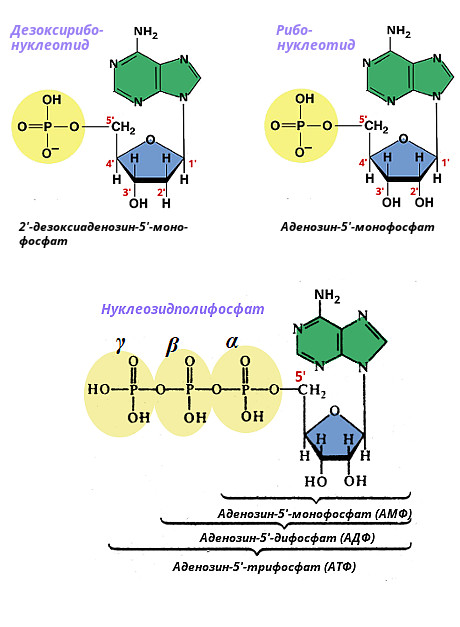
Фосфат может присоединяться в разные положения кольца пентозы (в рибонуклеотидах - в положениях 2", 3", 5", в дезоксирибонуклеотидах - в положения 3", 5"). Имеющиеся в клетке свободные нуклеотиды содержат фосфатную группу в положении 5". Нуклеозид-5"-фосфаты участвуют в биологическом синтезе нуклеиновых кислот и образуются при их распаде. Поскольку нуклеозид-5"-фосфаты, или мононуклеотиды, являются производными соответствующих нуклеозидов, то различают те же главные и редкие рибомононуклеотиды и дезоксирибомононуклеотиды.
Удлинение фосфатного конца мононуклеотида за счет присоединения дополнительных фосфатов приводит к образованию нуклеозидполифосфатов. Чаще всего в клетках встречаются нуклеозиддифосфаты и нуклеозидтрифосфаты. Ниже приводятся названия и сокращенные обозначения нуклеозидфосфатов:
Все нуклеозидфосфаты находятся в клетке в виде анионов, поэтому аденозинфосфаты правильнее обозначать АМФ 2- , АДФ 3- , АТФ 4- . АДФ и АТФ являются макроэргическими, т. е. богатыми энергией, соединениями, химическая энергия которых используется организмом для различных функций. Остальные нуклеозидди- и трифосфаты также участвуют в реакциях синтеза биологических веществ.
Международные стандартные сокращения
В работах по исследованию нуклеиновых кислот употребляются схемы нумерации атомов и сокращений, рекомендованные комиссией Международного союза общей и прикладной химии (IUPAC) и Международным союзом биохимиков (IUB). Подкомиссия IUPAC-IUB выработала единые стандартные определения (IUPAC-IUB, 1983).
Сокращения и символы, используемые для обозначения оснований, нуклеозидов и нуклеотидов (Arnott S., 1970).
| Основание | |||||
| Название | Символ | Название | Символ | Название | Символ |
| 1. Рибонуклеозиды и рибонуклеотиды | |||||
| Урацил | Ura | Уридин | Urd или U | Уридиловая кислота | 5"-UMP или pU |
| Цитозин | Cyt | Цитидин | Cyd или C | Цитидиловая кислота | 5"-CMP или pC |
| Аденин | Ade | Аденозин | Ado или A | Адениловая кислота | 5"-AMP или pA |
| Гуанин | Gua | Гуанозин | Guo или G | Гуаниловая кислота | 5"-GMP или pG |
| 2. Дезоксирибонуклеозиды и дезоксирибонуклеотиды | |||||
| Тимин | Thy | Дезокситимидин | dThd или dT | Дезокситимидиловая кислота | 5"-dTMP или pdT |
| Цитозин | Cyt | Дезоксицитидин | dCyd или dC | Дезоксицитидинловая кислота | 5"-dCMP или pdC |
| Аденин | Ade | Дезоксиаденозин | dAdo или dA | Дезоксиадениловая кислота | 5"dAMP или pdA |
| Гуанин | Gua | Дезоксигуанозин | dGuo или dG | Дезоксигуаниловая кислота | 5"dGMP или pdG |
| 3.Полинуклетиды | |||||
Синтетические полимеры, состоящие из нуклеотидов одного и того же типа, называют гомополимерами. Обозначение, например, полиадениловая кислота - poly(A) Синтетические полимеры с чередующейся последовательностью нуклеотидов называются гетерополимерами. Сополимер с чередованием dA и dT - poly(дезоксиаденилат - дезокситимидилат) обозначается как poly d(A-Т) или poly(dA-dT) или (dA-dT) или d(A-T)n. Для случайного сополимера dA, dT вместо деффиса между символами ставится запятая, например, poly d(A,T). Образование комплементарного дуплекса обозначается точкой между символами - poly(dA) · poly(dT); тройной спирали - poly(dA)· 2poly(dT). Олигонуклеотиды обозначаются следующим образом: например, олигонуклеотид гуанилил-3",5"-цитидилил-3",5"-уридин - GpCpU или GCU, при этом 5"-концевым нуклеотидом является G, а 3"-концевым - U. Для комплементарно связанных олигонуклеотидов номенклатура следующая: |
|||||

На рис.5. представлена принятая для нуклеотидов система нумерации атомов. Символы, обозначающие атомы сахара, отличаются от таковых для атомов оснований значком "штрих". Остов полинуклеотида описывают в направлении P -> O5" -> C5" -> C4" -> C3" -> O3" -> P.
В сахарном кольце нумерация такова: C1" -> C2" -> C3" -> C4" -> O4" ->C5".
Двум атомам водорода при атоме C5" и при атоме C2" в дезоксирибозе, а также двум свободным атомам кислорода при атомах фосфора приписываются номера 1 и 2, причем это делается следующим образом: если смотреть вдоль цепи в направлении O5"-> C5", то двигаясь по часовой стрелке, мы будем последовательно проходить атомы C4", H5"1, H5"2. Аналогично, если смотреть вдоль цепи в направлении O3" -> P - O5", то при движении по часовой стрелке мы будем последовательно проходить атомы O5", Op1, Op2.
Общая характеристика нуклеиновых кислот
Нуклеиновыми кислотами или полинуклеотидами называются высокомолекулярные вещества, состоящие из мононуклеотидов, соединенных в цепь 3",5"-фосфодиэфирными связями .
Общее содержание ДНК и РНК в клетках зависит от их функционального состояния. В сперматозоидах количество ДНК достигает 60% (в пересчете на сухую массу клеток), в большинстве клеток 1-10, а в мышцах около 0,2%. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в печени, поджелудочной железе, эмбриональных тканях и вообще в тканях, активно синтезирующих белок, составляет от 4 до 10. В тканях с умеренным синтезом белка соотношение колеблется от 0,3 до 2,5. Особое место занимают вирусы. У них в качестве генетического материала может быть либо ДНК (ДНК-овые вирусы), либо РНК (РНК-овые вирусы).
В клетках бактерий, не имеющих ядра (прокариоты), молекула ДНК (хромосома) находится в специальной зоне цитоплазмы - нуклеоиде. Если она связана с клеточной мембраной бактерии, то ее называют мезосомой. Фрагмент ДНК меньших размеров локализуется вне этой хромосомной зоны. Такие участки ДНК в бактериях называются плазмидами или эписомами. В клетках, имеющих ядро (эукариоты), ДНК распределена между ядром, где она входит в состав хромосом и ядрышка, и внеядерными органоидами (митохондриями и хлоропластами). Имеются наблюдения, что в очень малых количествах ДНК присутствует в микросомах.
Примерно 1-3% ДНК клетки приходится на внеядерную ДНК, а остальное сосредоточено в ядре. Значит, наследственные свойства характерны не только для ядра, но и для митохондрий и хлоропластов клеток. Необычно высоким содержанием внеядерной ДНК отличаются зрелые яйцеклетки, у которых она присутствует в многочисленных митохондриях и желточных пластинках, причем в последних является не генетическим материалом, а резервом нуклеотидов.
РНК в отличие от ДНК распределена по клетке более равномерно. Уже одно это обстоятельство говорит о том, что функция РНК более динамична и многообразна. В клетках высших организмов около 11% всей РНК находится в ядре, около 15% - в митохондриях, 50% - в рибосомах и 24% - в гиалоплазме.
Молекулярная масса ДНК зависит от степени сложности живого объекта: у бактерий она составляет 2 10 9 , у человека и животных достигает 10 11 . У бактерий ДНК находится в виде единичной гигантской молекулы, слабо связанной с белками. В других объектах ДНК окружена белками или простейшими аминами. У вирусов это простейшие основные белки или полиамины (путресцин и спермидин), которые нейтрализуют отрицательный заряд молекулы ДНК, связываясь с ее фосфатными группами. В сперматозоидах некоторых животных и рыб ДНК образует комплексы с протаминами и гистоноподобными белками. В хромосомах клеток человека и других высших организмов ДНК связана с гистонами и негистоновыми белками. Такие комплексы белка с ДНК называют дезоксирибонуклеопротеидами (ДНП).
РНК имеет значительно меньшую молекулярную массу, чем ДНК. В зависимости от выполняемой функции, молекулярной массы и состава нуклеотидов выделяют следующие главные типы РНК: информационная, или матричная (мРНК), транспортная (тРНК) и рибосомальная (рРНК). Разные рРНК различаются по молекулярной массе (табл. 13). Кроме трех главных типов есть минорные, или редкие, РНК, содержание которых в клетке незначительно, и функции их только изучаются.
Большинство типов РНК связано в клетке с различными белками. Такие комплексы называются рибонуклеопротеидами (РНП). Характеристика нуклеиновых кислот суммирована в табл. 1.
| Таблица 1. Краткая характеристика нуклеиновых кислот клеток высших организмов | |||||
| Тип нуклеиновой кислоты | Молекулярная масса | Константа седиментации (в единицах Сведберга-S) | Содержание в клетке, % | Локализация в клетке | Функция |
| ДНК | 10 11 | - | 97-99% от всей ДНК 1-3% от всей ДНК | Ядро Митохондрии | Хранение генетической информации и участие в передаче ее родительской ДНК при делении клетки или в передаче РНК в процессе жизнедеятельности |
| мРНК | 4 10 4 - 1,2 10 6 | 6-25 | 25% от всей РНК | Ядро, цитоплазма | Является копией участка ДНК, содержащего информацию о структуре полипептидной цепи белка. Переносит информацию от ДНК к месту синтеза белка - к рибосомам |
| тРНК | 2,5 10 4 | ~4 | 15% от всей РНК | Гиалоплазма, рибосомы, митохондрии | Участвует в активировании аминокислот, их транспорте к рибосомам и сборке из аминокислот полипептидов на рибосомах |
| рРНК | 0,7 10 6 | 18 | 80% от всей РНК | Рибосомы цитоплазмы | Образует скелет рибосом цитоплазмы (или митохондрий), который окутывается белками рибосом. Играет вспомогательную роль при сборке белка на рибосомах |
| 0,6 10 6 | 16 | Рибосомы митохондрий | |||
| ~4 10 4 | 5 | Все рибосомы | |||
| Хромосомная векторная РНК | 10 4 | 3 | Следы | Хромосомы ядер | Узнавание и активирование генов ДНК |
| Низкомолекулярные ядерные РНК | 2,5 10 4 -5 10 4 | 4-8 | Доли процента | Ядра, РНП частицы цитоплазмы | Активирование генов ДНК, формирование скелета белковых частиц, переносящих тРНК из ядра в цитоплазму |
Физико-химические свойства нуклеиновых кислот
Физико-химические свойства нуклеиновых кислот определяются высокой молекулярной массой и уровнем структурной организации. Для нуклеиновых кислот характерны: коллоидные и осмотические свойства, высокая вязкость и плотность растворов, оптические свойства, способность к денатурации.
Коллоидные свойства типичны для всех высокомолекулярных соединений. При растворении нуклеиновые кислоты набухают и образуют вязкие растворы типа коллоидов. Гидрофильность их зависит в основном от фосфатов. В растворе молекулы нуклеиновых кислот имеют вид полианиона с резко выраженными кислотными свойствами. При физиологических значениях pH все нуклеиновые кислоты являются полианионами и окружены противоионами из белков и неорганических катионов. Растворимость двуспиральных нуклеиновых кислот хуже, чем односпиральных.
Денатурация и ренатурация. Денатурация - свойство, присущее тем макромолекулам, которые имеют пространственную организацию. Денатурация вызывается нагреванием, воздействием химических веществ, которые разрывают водородные и ван-дер-ваальсовы связи, стабилизирующие вторичную и третичную структуру нуклеиновых кислот. Например, нагревание ДНК приводит к разделению двойной спирали на одиночные цепи, т. е. наблюдается переход "спираль - клубок". При медленном охлаждении цепи вновь воссоединяются по принципу комплементарности. Образуется нативная двойная спираль ДНК. Это явление называется ренатурацией. При быстром охлаждении ренатурация не происходит.
Характерно изменение оптической активности нуклеиновых кислот, сопровождающее их денатурацию и ренатурацию. Спиральные (организованные) участки нуклеиновых кислот вращают плоскость поляризованного света, т. е. оптически активны, а разрушение спиральных участков сводит на нет оптическую активность нуклеиновых кислот.
Все нуклеиновые кислоты имеют максимум оптической плотности при длине волны около 260 нм, что соответствует максимуму поглощения азотистых оснований. Однако интенсивность поглощения природной нуклеиновой кислоты значительно ниже, чем смеси ее же нуклеотидов, полученных, например, при гидролизе этой нуклеиновой кислоты, или одиночных цепей. Причиной является структурная организация ДНК и РНК, которая вызывает классический эффект - снижение оптической плотности. Это явление получило название гипохромного эффекта. Он максимально выражен у нуклеиновых кислот, имеющих спиральные структуры (например, у ДНК) и содержащих много ГЦ-пар (ГЦ-пары имеют три водородные связи, и поэтому их труднее разорвать).
Молекулярная гибридизация нуклеиновых кислот. На способности нуклеиновых кислот ренатурировать после денатурации основан чрезвычайно важный метод определения степени гомологичности, или родственности, нуклеиновых кислот. Его называют молекулярной гибридизацией. В его основе лежит комплементарное спаривание одноцепочечных участков нуклеиновых кислот.
Этот метод позволил обнаружить особенности первичной структуры ДНК. Оказывается, в ДНК животных имеются многократно (до 100 000 раз) повторяющиеся участки с одинаковой последовательностью нуклеотидов. Они составляют до 10-20% всей ДНК. Их гибридизация идет очень быстро. Остальная часть ДНК представлена уникальными последовательностями, которые не дублируются. Эти участки ДНК гибридизуются очень медленно. Вероятность их совпадения у разных организмов невелика. С помощью метода молекулярной гибридизации можно установить гомологичность ДНК организма одного вида ДНК другого вида или гомологичность РНК участкам ДНК.
Нуклеиновые кислоты и систематика организмов
Нуклеиновые кислоты являются материальным носителем наследственной информации и определяют видоспецифичность организма, сложившуюся в ходе эволюции. Изучение особенностей нуклеотидного состава ДНК разных организмов позволило перейти от систематики по внешним признакам к систематике генетической. Это направление в молекулярной биологии получило название геносистематики. Основателем его был выдающийся советский биохимик А. Н. Белозерский.
Сравнение нуклеотидного состава ДНК разных организмов привело к интересным выводам. Оказалось, что коэффициент специфичности ДНК, т. е. отношение Г + Ц к А + Т, сильно варьирует у микроорганизмов и довольно постоянен у высших растений и животных. У микроорганизмов наблюдаются колебания изменчивости от крайнего ГЦ-типа до выраженного АТ-типа. ДНК высших организмов стойко сохраняет АТ-тип. Может создаться впечатление, что у высших организмов теряется специфичность ДНК. На самом деле у них она так же специфична, как и у бактерий, но ее специфичность определяется не столько изменчивостью состава нуклеотидов, сколько последовательностью чередования их вдоль цепи. Интересные выводы на основании нуклеотидного состава ДНК были сделаны А. Н. Белозерским и его учениками относительно происхождения многоклеточных животных и высших растений. Их ДНК АТ-типа ближе всего к ДНК грибов, поэтому свою родословную животные и грибы, очевидно, ведут от общего предка - крайне примитивных грибообразных организмов.
Еще большую информацию о родстве организмов дает метод молекулярной гибридизации. С помощью этого метода была установлена высокая гомологичность ДНК человека и обезьяны. Причем по составу ДНК человека всего на 2-3% отличается от ДНК шимпанзе, чуть больше - от ДНК гориллы, более чем на 10% - от ДНК остальных обезьян, а от ДНК бактерии - почти на 100%. Особенности первичной структуры ДНК тоже можно использовать в систематике. Гомология по участкам повторяющихся последовательностей (быстрая гибридизация) используется для макросистематики, а для уникальных фрагментов ДНК (медленная гибридизация) - для микросистематики (на уровне видов и родов). Ученые считают, что постепенно по ДНК можно будет построить все родословное древо живого мира.
К 1944 г. О. Эйвери и его коллеги К. Маклеод и М. Маккарти открыли трансформирующую активность ДНК у пневмококков. Эти авторы продолжили работу Гриффита, описавшего феномен трансформации (передачи наследственных признаков) у бактерий. О. Эйвери, К. Маклеод, М. Маккарти показали, что при удалении белков, полисахаридов и РНК трансформация бактерий не нарушается, а при воздействии на индуцирующее вещество ферментом дезоксирибонуклеазой трансформирующая активность исчезает.
В этих экспериментах впервые была продемонстрирована генетическая роль молекулы ДНК. В 1952 г. А. Херши и М. Чейз подтвердили генетическую роль молекулы ДН К в опытах на бактериофаге Т2. Пометив его белок радиоактивной серой, а ДНК-радиоактивным фосфором,они инфицировали этим бактериальным вирусом кишечную палочку Е. coli. В потомстве фага было выявлено большое количество радиоактивного фосфора и лишь следы S. Отсюда следовало, что именно ДНК, а не белок фага проникает в бактерию, а затем после репликации передается фаговому потомству.
Строение нуклеотида днк. Типы нуклеотидов.
Нуклеотид ДНК состоит из
Азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
Моносахара дезоксирибозы
Фосфорной кислоты
Молекула нуклеотида состоит из трех частей - пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар, входящий в состав нуклеотида , содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два типа нуклеиновых кислот - рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу. В дезоксирибозе - ОН-группа при 2-м атоме углерода заменена на атом Н, т. е. в ней на один атом кислорода меньше, чем в рибозе.
В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: два из них относятся к классу пуринов и два - к классу пиримидинов. Основной характер этим соединениям придает включенный в кольцо азот. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов - цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК). Тимин химически очень близок к урацилу (он представляет собой 5-метилурацил, т. е. урацил, в котором у 5-го углеродного атома стоит метильная группа). В молекуле пуринов имеется два кольца, а в молекуле пиримидинов - одно.
Нуклеотиды соединяются между собой прочной ковалентной связью через сахар одного нуклеотида и фосфорную кислоту другого. Получаетсяполинуклеотидная цепь . На одном ее конце – свободная фосфорная кислота (5’-конец), на другом – свободный сахар (3’-конец). (ДНК-полимераза может присоединять новые нуклеотиды только к 3’-концу.)
Две полинуклеотидные цепи соединяются друг с другом слабыми водородными связями между азотистыми основаниями. Соблюдаются 2 правила:
принцип комплементарности: напротив аденина всегда стоит тимин, напротив цитозина – гуанин (они подходят друг другу по форме и числу водородных связей – между А и Г две связи, между Ц и Г – 3).
принцип антипараллельности: там, где у одной полинуклеотидной цепи 5’-конец, у другой – 3’-конец, и наоборот.
Получается двойная цепь ДНК.
Она скручивается в двойную спираль , один виток спирали имеет длину 3,4 нм, содержит 10 пар нуклеотидов. Азотистые основания (хранители генетической информации) находятся внутри спирали, защищенные.
Все живое на планете состоит из множества клеток, поддерживающих упорядоченность своей организации за счет содержащейся в ядре генетической информации. Она сохраняется, реализуется и передается сложными высокомолекулярными соединениями - нуклеиновыми кислотами, состоящими из мономерных звеньев - нуклеотидов. Роль нуклеиновых кислот невозможно переоценить. Стабильностью их структуры определяется нормальная жизнедеятельность организма, а любые отклонения в строении неминуемо приводят к изменению клеточной организации, активности физиологических процессов и жизнеспособности клеток в целом.
Понятие нуклеотида и его свойства
Каждая или РНК собрана из более мелких мономерных соединений - нуклеотидов. Другими словами, нуклеотид - это строительный материал для нуклеиновых кислот, коферментов и многих других биологических соединений, которые крайне необходимы клетке в процессе ее жизнедеятельности.
К основным свойствам этих незаменимых веществ можно отнести:
Хранение информации о и наследуемых признаках;
. осуществление контроля над ростом и репродукцией;
. участие в метаболизме и многих других физиологических процессах, протекающих в клетке.
Говоря о нуклеотидах, нельзя не остановиться на таком важном вопросе, как их структура и состав.

Каждый нуклеотид состоит из:
Сахарного остатка;
. азотистого основания;
. фосфатной группы или остатка фосфорной кислоты.
Можно сказать, что нуклеотид - это сложное органическое соединение. В зависимости от видового состава азотистых оснований и типа пентозы в структуре нуклеотида нуклеиновые кислоты подразделяются на:
Дезоксирибонуклеиновую кислоту, или ДНК;
. рибонуклеиновую кислоту, или РНК.
Состав нуклеиновых кислот
В нуклеиновых кислотах сахар представлен пентозой. Это пятиуглеродный сахар, в ДНК его называют дезоксирибозой, в РНК - рибозой. Каждая молекула пентозы имеет пять атомов углерода, четыре из них вместе с атомом кислорода образуют пятичленное кольцо, а пятый входит в группу НО-СН2.
Положение каждого атома углерода в молекуле пентозы обозначается арабской цифрой со штрихом (1C´, 2C´, 3C´, 4C´, 5C´). Поскольку все процессы считывания с молекулы нуклеиновой кислоты имеют строгую направленность, нумерация атомов углерода и их расположение в кольце служат своего рода указателем правильного направления.
По гидроксильной группе к третьему и пятому углеродным атомам (3С´ и 5С´) присоединен остаток фосфорной кислоты. Он и определяет химическую принадлежность ДНК и РНК к группе кислот.
К первому углеродному атому (1С´) в молекуле сахара присоединено азотистое основание.
Видовой состав азотистых оснований
Нуклеотиды ДНК по азотистому основанию представлены четырьмя видами:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. тимином (Т).
Первые два относятся к классу пуринов, два последних - пиримидинов. По молекулярной массе пуриновые всегда тяжелее пиримидиновых.

Нуклеотиды РНК по азотистому основанию представлены:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. урацилом (У).
Урацил так же, как и тимин, является пиримидиновым основанием.
В научной литературе нередко можно встретить и другое обозначение азотистых оснований - латинскими буквами (A, T, C, G, U).
Подробнее остановимся на химической структуре пуринов и пиримидинов.

Пиримидины, а именно цитозин, тимин и урацил, в своем составе представлены двумя атомами азота и четырьмя атомами углерода, образующих шестичленное кольцо. Каждый атом имеет свой номер от 1 до 6.
Пурины (аденин и гуанин) состоят из пиримидина и имидазола или двух гетероциклов. Молекула пуриновых оснований представлена четырьмя атомами азота и пятью атомами углерода. Каждый атом пронумерован от 1 до 9.
В результате соединения азотистого основания и остатка пентозы образуется нуклеозид. Нуклеотид - это соединение нуклеозида и фосфатной группы.
Образование фосфодиэфирных связей
Важно разобраться в вопросе о том, как соединяются нуклеотиды в полипептидную цепь и образуют молекулу нуклеиновой кислоты. Происходит это за счет так называемых фосфодиэфирных связей.
Взаимодействие двух нуклеотидов дает динуклеотид. Образование нового соединения происходит путем конденсации, когда между фосфатным остатком одного мономера и гидроксигруппой пентозы другого возникает фосфодиэфирная связь.
Синтез полинуклеотида - неоднократное повторение этой реакции (несколько миллионов раз). Полинуклеотидная цепь строится посредством образования фосфодиэфирных связей между третьим и пятым углеродами сахаров (3С´ и 5С´).
Сборка полинуклеотида - сложный процесс, протекающий при участии фермента ДНК-полимеразы, которая обеспечивает рост цепи только с одного конца (3´) со свободной гидроксигруппой.
Структура молекулы ДНК
Молекула ДНК, так же как и белка, может иметь первичную, вторичную и третичную структуру.

Последовательность нуклеотидов в цепи ДНК определяет ее первичную формируется за счет водородных связей, в основе возникновения которых положен принцип комплементарности. Другими словами, при синтезе двойной действует определенная закономерность: аденин одной цепи соответствует тимину другой, гуанин - цитозину, и наоборот. Пары аденина и тимина или гуанина и цитозина образуются за счет двух в первом и трех в последнем случае водородных связей. Такое соединение нуклеотидов обеспечивает прочную связь цепей и равное расстояние между ними.
Зная последовательность нуклеотидов одной цепи ДНК, по принципу комплементарности или дополнения можно достроить вторую.
Третичная структура ДНК образована за счет сложных трехмерных связей, что делает ее молекулу более компактной и способной размещаться в малом объеме клетки. Так, например, длина ДНК кишечной палочки составляет более 1 мм, тогда как длина клетки - меньше 5 мкм.
Число нуклеотидов в ДНК, а именно их количественное соотношение, подчиняется правилу Чергаффа (число пуриновых оснований всегда равно количеству пиримидиновых). Расстояние между нуклеотидами - величина постоянная, равная 0,34 нм, как и их молекулярная масса.
Структура молекулы РНК
РНК представлена одной полинуклеотидной цепочкой, образованной через между пентозой (в данном случае рибозой) и фосфатным остатком. По длине она значительно короче ДНК. По видовому составу азотистых оснований в нуклеотиде также имеются различия. В РНК вместо пиримидинового основания тимина используется урацил. В зависимости от функций, выполняемых в организме, РНК может быть трех типов.

Рибосомальная (рРНК) - содержит обычно от 3000 до 5000 нуклеотидов. Как необходимый структурный компонент принимает участие в формировании активного центра рибосом, места осуществления одного из важнейших процессов в клетке — биосинтеза белка.
. Транспортная (тРНК) - состоит в среднем из 75 - 95 нуклеотидов, осуществляет перенос нужной аминокислоты к месту синтеза полипептида в рибосоме. Каждый вид тРНК (не менее 40) имеет свою, присущую только ему последовательность мономеров или нуклеотидов.
. Информационная (иРНК) - по нуклеотидному составу весьма разнообразна. Переносит генетическую информацию от ДНК к рибосомам, выступает в роли матрицы для синтеза белковой молекулы.
Роль нуклеотидов в организме
Нуклеотиды в клетке выполняют ряд важнейших функций:
Используются в качестве структурных блоков для нуклеиновых кислот (нуклеотиды пуринового и пиримидинового рядов);
. участвуют во многих обменных процессах в клетке;
. входят в состав АТФ - главного источника энергии в клетках;
. выступают в роли переносчиков восстановительных эквивалентов в клетках (НАД+, НАДФ+, ФАД, ФМН);
. выполняют функцию биорегуляторов;
. могут рассматриваться как вторые вестники внеклеточного регулярного синтеза (например, цАМФ или цГМФ).
Нуклеотид - это мономерная единица, образующая более сложные соединения - нуклеиновые кислоты, без которых невозможна передача генетической информации, ее хранение и воспроизведение. Свободные нуклеотиды являются главными компонентами, участвующими в сигнальных и энергетических процессах, поддерживающих нормальную жизнедеятельность клеток и организма в целом.
4.2.1. Первичной структурой нуклеиновых кислот называется последовательность расположения мононуклеотидов в цепи ДНК или РНК . Первичная структура нуклеиновых кислот стабилизируется 3",5"-фосфодиэфирными связями. Эти связи образуются при взаимодействии гидроксильной группы в 3"-положении пентозного остатка каждого нуклеотида с фосфатной группой соседнего нуклеотида (рисунок 3.2),
Таким образом, на одном конце полинуклеотидной цепи имеется свободная 5"-фосфатная группа (5"-конец), а на другом - свободная гидроксильная группа в 3"-положении (3"-конец). Нуклеотидные последовательности принято записывать в направлении от 5"-конца к 3"-концу.
Рисунок 4.2. Структура динуклеотида, в состав которого входят аденозин-5"-монофосфат и цитидин-5"-монофосфат.
4.2.2. ДНК (дезоксирибонуклеиновая кислота) содержится в клеточном ядре и имеет молекулярную массу порядка 1011 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, тимин , углевод дезоксирибоза и остатки фосфорной кислоты. Содержание азотистых оснований в молекуле ДНК определяют правила Чаргаффа:
1) количество пуриновых оснований равно количеству пиримидиновых (А + Г = Ц + Т) ;
2) количество аденина и цитозина равно количеству тимина и гуанина соответственно (А = Т; Ц = Г) ;
3) ДНК, выделенные из клеток различных биологических видов, отличаются друг от друга величиной коэффициента специфичности:
(Г + Ц) /(А + Т)
Эти закономерности в строении ДНК объясняются следующими особенностями её вторичной структуры:
1) молекула ДНК построена из двух полинуклеотидных цепей, связанных между собой водородными связями и ориентированных антипараллельно (то есть 3"-конец одной цепи расположен напротив 5"-конца другой цепи и наоборот);
2) водородные связи образуются между комплементарными парами азотистых оснований. Аденину комплементарен тимин; эта пара стабилизируется двумя водородными связями. Гуанину комплементарен цитозин; эта пара стабилизируется тремя водородными связями (см. рисунок б) . Чем больше в молекуле ДНК пар Г-Ц, тем больше её устойчивость к действию высоких температур и ионизирующего излучения;

Рисунок 3.3. Водородные связи между комплементарными азотистыми основаниями.
3) обе цепи ДНК закручены в спираль, имеющую общую ось. Азотистые основания обращены внутрь спирали; кроме водородных, между ними возникают также гидрофобные взаимодействия. Рибозофосфатные части расположены по периферии, образуя остов спирали (см. рисунок 3.4).

Рисунок 3.4. Схема строения ДНК.
4.2.3. РНК (рибонуклеиновая кислота) содержится преимущественно в цитоплазме клетки и имеет молекулярную массу в пределах 104 - 106 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, урацил , углевод рибоза и остатки фосфорной кислоты. В отличие от ДНК, молекулы РНК построены из одной полинуклеотидной цепи, в которой могут находиться комплементарные друг другу участки (рисунок 3.5). Эти участки могут взаимодействовать между собой, образуя двойные спирали, чередующиеся с неспирализованными участками.

Рисунок 3.5. Схема строения транспортной РНК.
По особенностям структуры и функции различают три основных типа РНК:
1) матричные (информационные) РНК (мРНК) передают информацию о структуре белка из клеточного ядра на рибосомы;
2) транспортные РНК (тРНК) осуществляют транспорт аминокислот к месту синтеза белка;
3) рибосомальные РНК (рРНК) входят в состав рибосом, участвуют в синтезе белка.